|
|
|
1. 序 生物の生存には、遺伝子に保存されている情報を書き出す必要があります。これが遺伝子発現です。生命活動に必要とされるタンパク質やRNAを遺伝情報に基づいて合成することです。具体的にはどのようなことが生じているのでしょうか? ここでは、遺伝子発現に関する反応を真核生物について分子レベルで説明していきたいと思います。
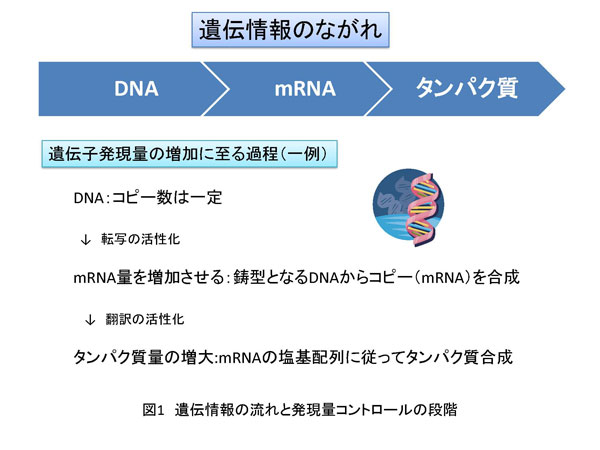
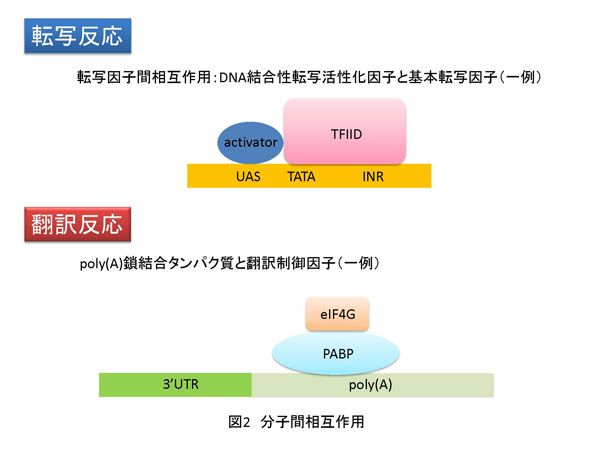
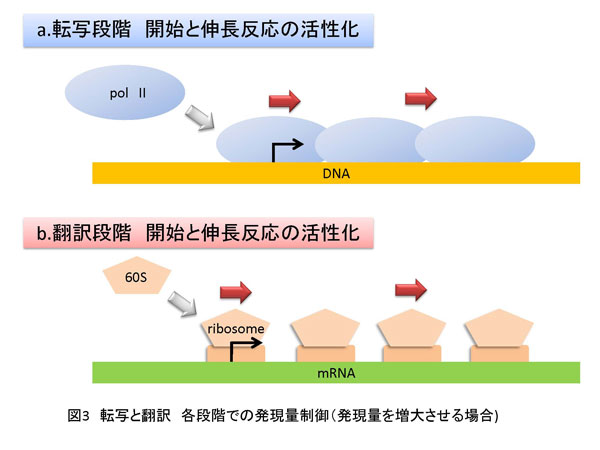
2. 遺伝子発現過程と発現量のコントロール 遺伝情報は核という構造体中のゲノムDNAに塩基配列情報として保存されています。核は、核膜によって細胞質と区別されます。ゲノムDNA上の特定の領域はRNA重合酵素(ポリメラーゼ)により転写され、DNAの塩基配列情報のコピーとなるmRNAが合成されます。合成されたmRNAは核から抜け出し細胞質に存在するリボソームに移動します。リボソームは翻訳反応によりmRNA上の塩基配列情報に従ってタンパク質を合成します。その後、タンパク質は翻訳後修飾と呼ばれる化学修飾を受けて本来の機能を発揮する状態になります。このような素反応が連続して展開される一連の反応を遺伝子発現過程と呼び、DNA→mRNA→タンパク質という情報の流れをセントラルドグマと言います。この過程において重要な制御の一つに遺伝子の発現量コントロールがあります(図1)。遺伝子発現量は、転写や翻訳効率、mRNAやタンパク質の安定性、あるいはmRNAの核から細胞質への移動速度、さらにこれら以外にもさまざまな段階で制御されています。ただし、このような異なる各ステップにおいて共通していることがあります。それは、それぞれの遺伝子発現にかかわる素反応は、分子(核酸−核酸、核酸−タンパク質およびタンパク質-タンパク質)間相互作用に基づいているということです(図2)。また、核やリボソームといった細胞小器官(オルガネラ)や細胞に特徴的な構造体、例えば核マトリックスなどが重要な役割を持つことも忘れてはなりません。次は、遺伝子発現過程の中で特に転写反応と翻訳反応段階での制御に関して説明したいと思います。 2-1. 転写反応における遺伝子発現量制御-mRNA合成効率のコントロール 転写反応はRNAポリメラーゼII(pol II)によるmRNA合成反応のことで、核内で生じます。転写反応は大きく3つのステップ、すなわち 開始 伸長 終結 に分けることができます。開始反応は、転写反応の鋳型となるゲノムDNA上の特定遺伝子の転写制御領域(プロモーター)にpol II(上記RNA重合酵素の一種)が結合し、転写開始点で2つのヌクレオチドをつなげてリン酸ジエステル結合を作ることです。伸長反応ではそれぞれのヌクレオチドがDNAの塩基配列に従って規則正しく重合し、ポリヌクレオチド鎖が伸長します。終結反応は文字通り転写反応を終わらせる反応です。開始反応の詳細やそれに至るまでの段階の制御は、古久保先生の説明をご参照ください。伸長反応における制御に関しては、後ほど詳しく説明します。終結反応ついては、教科書等で学んでいただくとしてここでは説明いたしません。 翻訳反応においても転写反応と同様に反応は大きく分けて 開始 伸長 終結 の3つのステップからなり、翻訳活性化とは1つのmRNAからたくさんのタンパク質が短時間に合成されることを意味します。もう少し具体的に説明するならば、リボソームがmRNA上の翻訳開始コドン(ATG)からポリペプチド合成を開始する頻度が上昇することやポリペプチド鎖伸長効率が上昇することです(図3b)。
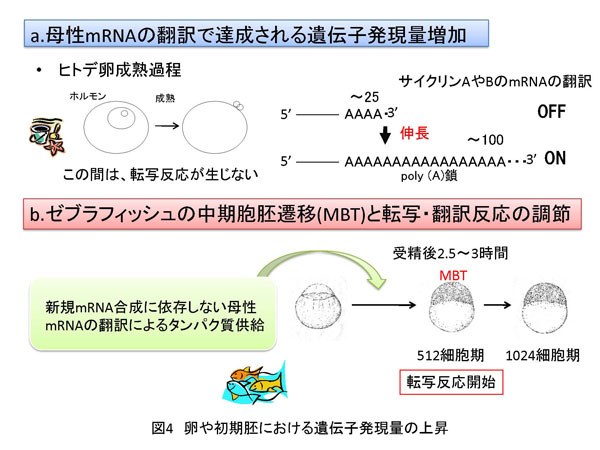
さて、ここで話をこれまでとは全く別の方向に進めたいと思います。卵は雌側の配偶子のことです。対応するのが雄の精子です。卵と精子が結合(受精)することによりDNA量が倍加して、細胞分裂(卵割)が始まります。私は、最近、実験材料として色々な動物の卵を取り扱うようになりました。第一の理由は、細胞周期が同調しているからです。第二の理由は、転写反応が一時的に停止しているからです。詳細は以下に示します。 最初にお断りしておきますが、卵の成熟過程の詳細に関しては、他の専門書や教科書をご参照ください。ここでは、転写や翻訳反応を解析するために必要な知識に絞って紹介します。一般に卵は成熟ホルモンの刺激を受けて2回の連続した減数分裂を経験します。その間に、極体が放出されます。その後、細胞周期は停止します。停止時期は動物の種類により様々です。私が興味深く思っていることは、卵成熟中にほとんど、いや全くと言ってよいほど転写反応が生じないということです。別の言い方をします。成熟ホルモンの刺激後の減数分裂過程やそれ以降の受精、さらには初期発生段階で必要となるタンパク質の供給は、卵細胞の細胞質に蓄積されているいわゆる母性(maternal)mRNAが翻訳されることによって供給され、新規のmRNA合成には依存しません(図4a)。まとめると、卵成熟過程の遺伝子発現は、mRNA合成を伴わない母性mRNAの翻訳でまかなわれると言うことです。 受精では成熟した卵に精子が結合してDNA量が倍加し、この後卵割が始まります。私の研究材料の一つの小型熱帯魚ゼブラフィッシュでは、興味深いことに通常の細胞分裂と比較すると極めて早い速度で初期の卵割が進行します。この速さは、細胞周期におけるギャップ期、すなわちG1期やG2期が存在せずに卵割が進行することに他なりません。これに関しても、詳細は他の教科書等をご参照ください。もう一点、初期発生で興味深い点は、卵成熟過程と同様に、ある程度の分裂回数まで転写がほとんど起こらないということです。ゼブラフィッシュでは、受精後2時間半から3時間頃まで、分裂回数で言うと9回目から10回目にかけてまで転写反応は抑制された状態を維持しています(図4b)。よって、この段階でも新規mRNA合成に依存しない母性mRNAの翻訳によるタンパク質供給で遺伝子発現は生じています。
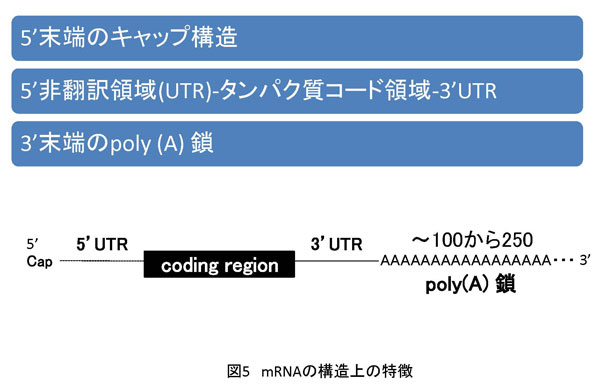
4. 卵および初期胚の母性mRNAの構造的特徴 母性mRNAの構造に関して言及する前に、一般的なmRNAの構造から説明したいと思います。mRNAは5'側にキャップ構造、それに続く5'非翻訳領域(UnTranslated Region;5'UTR)、次いでコーディング領域(翻訳領域、タンパク質の一次構造を決めるための塩基配列)、そして3'UTR、さらに3'末端の「A」が連続して重合しているいわゆるpoly(A)鎖が存在します(図5)。 卵成熟中は転写反応がほとんど停止しているので、mRNAの提供なしに遺伝子発現が生じます。それでは、どのようにして卵細胞質中に蓄積している母性mRNAの翻訳反応が促進されるのでしょうか?それは、先に説明したpoly(A)鎖長変化と密接に関係します。簡単に説明するならば、短鎖状態では翻訳反応が抑制されていて、長鎖になると活性化されるということです(図6)。この制御にはmRNAの3'UTR内の特異塩基配列とそこに結合するタンパク質群によって形成される高次複合体による翻訳活性のコントロールが密接に関係しています。卵成熟ホルモンの暴露で卵細胞のシグナル伝達系が刺激され、その結果3'UTRにおける複合体が変化することでpoly(A)鎖が伸長し、さらにこれが複合体の変化を促して最終的には翻訳反応の活性化に至る経路が予想されています。複合体を形成する因子名や詳しい分子メカニズムに関しては、私の研究がもう少し進展したら随時紹介していくことにします。 卵と同様に、初期胚においてもpoly(A)鎖伸長と翻訳反応の活性化の関係が成立します。私はゼブラフィッシュのCdk9のmRNAに関して母性mRNAのpoly(A)鎖伸長は受精後およそ2時間くらいから開始し、4時間後に最長となり、その後短小化することを見出しています。また、その鎖長変化に少し遅れるようにして、翻訳の活性化、すなわちCdk9タンパク質の蓄積量の増加を観察することができます。ここで私はCdk9をコードする母性mRNAのpoly(A)鎖の伸長が本当に翻訳反応の活性化に寄与するのか解析する目的で、天然には存在しない核酸類似体でCdk9母性mRNAのpoly(A)鎖伸長の特異的阻害を試してみました。その結果、poly(A)鎖の伸長が阻害されるとCdk9タンパク質蓄積量の増加が抑制されることを見出しました。よって、ゼブラフィッシュ初期胚のCdk9母性mRNAは、poly(A)鎖の伸長でその発現量が増加することを明らかにしたわけです。
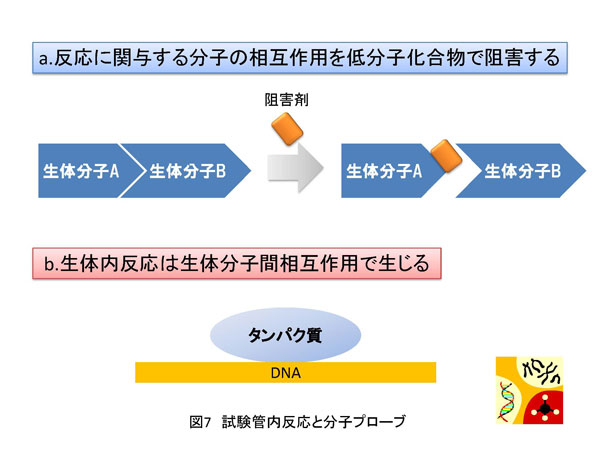
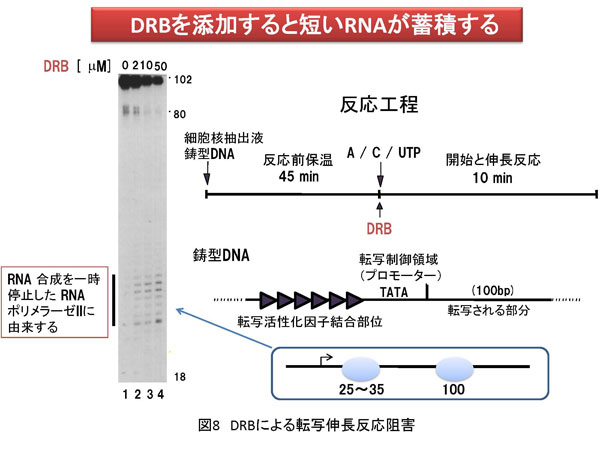
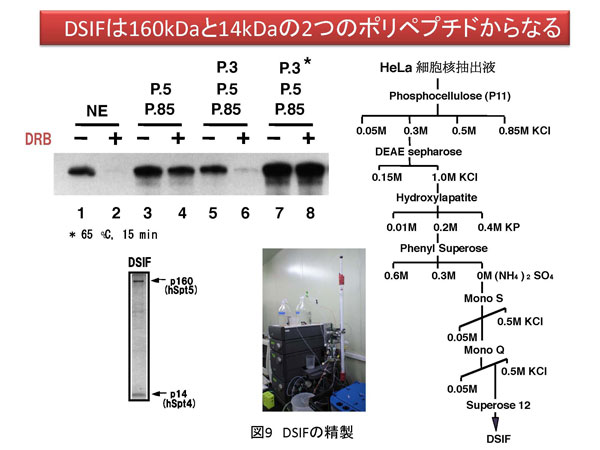
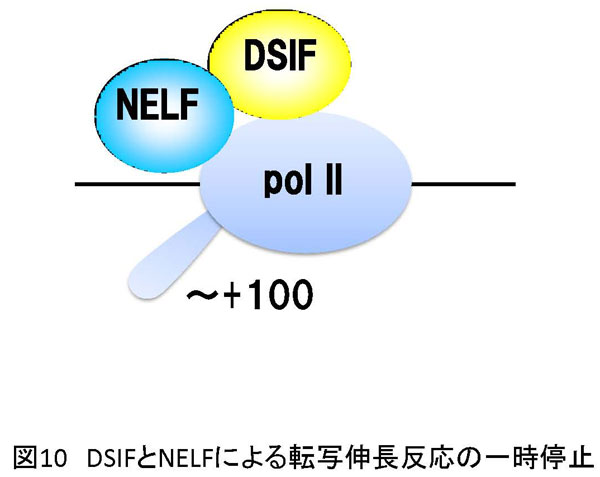
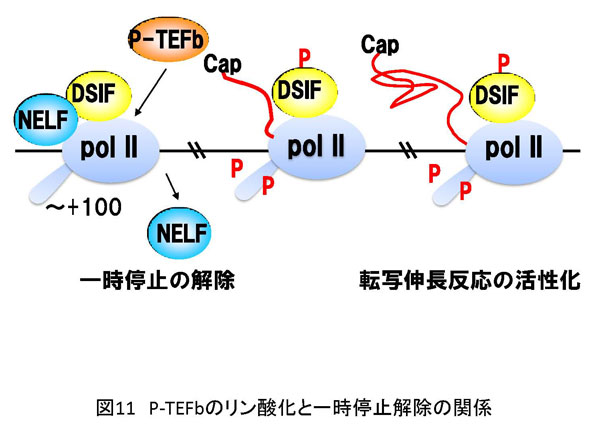
5. 方法論 生命現象に関わる反応を解析する方法は多数存在します。その中で、私は、生体内反応の分子レベルでの理解を目的として、生化学的アプローチと分子生物学的アプローチを組み合わせた方法を採用しています。特に私は、試験管内再構成系を用いた実験手法にこだわりがあります。以下に詳しく説明しますが、研究対象とする反応を分子レベルで解析する目的で低分子化合物(化学物質)を実験用ツール(分子プローブ)として使用しています。試験管内で再構成した生体内反応を分子プローブで阻害することにより解析の糸口を見出していこうとする試みです。これが私の研究の特徴と言えます(図7a)。簡単に説明するならば、阻害剤で平衡を乱すことで生じる変化から、反応制御に密接に関わる因子を同定し、その因子の機能を明らかにすることで生体内反応の分子レベルでの理解を目指しています。多くの場合、阻害剤は核酸あるいはタンパク質と特異的に結合することで結合相手の本来の活性に変化を与えます。最初の項で説明しましたが、諸々の生体内反応の基本は分子間相互作用です(図7b)。よって、阻害剤が標的分子に結合することでこの分子間結合が切れたり、弱くなったり、あるいは増強されることにより平衡の乱れを誘導します。試験管内再構成系では限られた分子で反応が再現されていますので、その反応系が高度に精製された因子群から再構成されている場合は、阻害剤の標的因子、すなわち結合する相手探しが極めて容易となるわけです。同定された因子の機能や活性は、遺伝子ノックダウン法により細胞あるいは実験動物で解析を行うことが最近のサイエンスでは可能となりました。まとめると、[試験管内再構成系]→[阻害剤の作用メカニズムの解析]→[責任因子の同定]→[阻害メカニズムの解析]→[細胞あるいは個体レベルでの検証] という実験過程を辿るわけです。 私の阻害剤を用いた転写制御研究結果に関してここで紹介したいと思います。採用した阻害剤はATP類似体で、タンパク質リン酸化酵素(キナーゼ)の活性を特異的に阻害するDRBという低分子化合物です。世界で最初のDRBに関する報告は1950年代で、その後私が研究を開始した1995年頃までに、多くの論文にDRBに関する記述が存在しました。その中で、当時、私が特に重要と認識していた現象は「DRBはある種のキナーゼ活性を阻害することにより転写反応の伸長段階を阻害する」というものでした。「ある種の」と表記した理由は、DRBの標的が不明であったことに他なりません。 分子プローブとして採用したDRB研究から、転写伸長反応の制御を司るDSIFとNELFの同定に至ったわけです。しかし、それら因子を同定しただけではDRBによる転写伸長反応メカニズムを明らかにしたわけではありません。これを達成するために、私たちはin vitro転写反応を再構成系で行うことによりDRB阻害機構を解析することにしました。再構成系とは、できるだけ精製した転写因子や組換えタンパク質を用いて転写反応を実施することです。これにより、DRB標的因子の同定が容易となります。結果として、我々は米国David Price博士らのグループがショウジョウバエ細胞抽出液を用いて世界で最初に同定したキナーゼP-TEFbがDRBの標的であることを突き止めました。P-TEFbがリン酸化するものはpol IIに特徴的な構造のC-terminal domain(CTD)のセリン残基です。CTDは7つのアミノ酸から構成されているのですが、P-TEFbは2番目に位置するセリン残基を特異的にリン酸化することがわかりました。このリン酸化によりDSIFとNELFはpol IIと結合できなくなり、その結果転写伸長反応の一時停止は生じなくなります。すなわち、DRBがP-TEFb活性を阻害するとCTDのリン酸化は抑制され、DSIFとNELFがpol IIに結合した状態が維持されることにより転写反応の一時停止が持続するというDRB阻害分子メカニズムの詳細が明らかとなりました(図11)。 反応阻害剤、すなわち分子プローブは標的分子との結合を介して化学反応に影響を与えます。一方で、試験管内再構成系は、高度に精製した因子や組換えタンパク質で組み立てることが可能です。よって、先ほど説明したDRB研究のように、反応制御過程やそれに関与する因子の同定が可能となるわけです。すなわち、両者の組み合わせは約束された成果を与える有益な実験系であると考えられます。 先に紹介しましたが、私は最近、ゼブラフィッシュのCdk9をコードする母性mRNAを研究対象として、初期胚中でpoly(A)鎖伸長を特異的に阻害すると翻訳反応の促進が生じなくなることを発見しました。同様の現象は、イトマキヒトデのサイクリンAやBをコードする母性mRNAでも確認しています。さらに、アフリカツメガエル卵においても検証中です。もし、各遺伝子のmRNAのpoly(A)鎖伸長を抑制する低分子化合物を発見できれば、これを応用することにより特異的遺伝子発現抑制法に展開させることが可能となります。また、転写阻害剤DRBの実験例で説明しましたように、試験管内で反応を再構成することで反応の分子メカニズムを明らかにできます。よって、翻訳反応やpoly(A)鎖伸長反応を試験管内で再現し、この系でpoly(A)鎖伸長を特異的に阻害する化合物の活性を検討できれば責任分子の同定や解析の道が開けてきます。幸いアフリカツメガエル卵抽出液では、1)poly(A)鎖伸長と翻訳の活性化の関係が試験管内で解析されている、2)部分的ではあるが翻訳反応の再構成系の成功例が報告されている、などの状況が整っています。今後は、アフリカツメガエル卵抽出液を用いてpoly(A)鎖伸長阻害剤の作用メカニズムの解析を中心に研究を展開していく予定です。
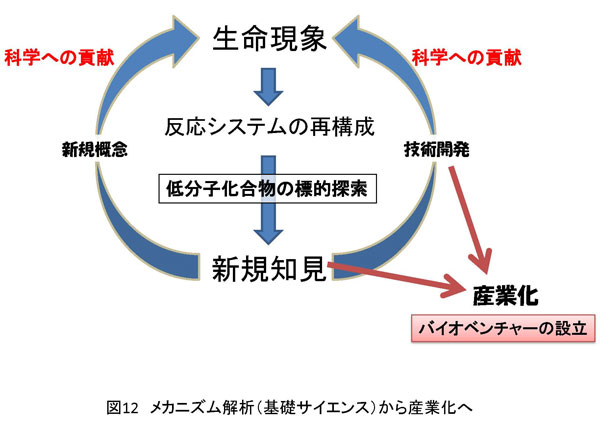
6. 技術の応用 分子レベルで生体内反応を理解すると、目的の反応において重要な分子間相互作用が明らかになります。そのことはすなわち、反応の活性変化を特異的に誘導できる箇所がわかるということです。もう少し具体的に説明するならば、反応の活性に重要な分子間相互作用を阻害する低分子化合物の探索が可能となるわけです(図12)。そうすることで、ある種の薬剤の開発や遺伝子発現制御技術の発展が見込めます。 よって、目的の分子間相互作用を変化させる低分子化合物を特定の遺伝子やその遺伝子産物に絞り創製したならば、それは新規の遺伝子発現抑制法に至る経路の確立へとつながります。 もし、遺伝子特異的発現抑制法が確立できたならば、例えばがん細胞の増殖に必須な遺伝子発現を抑制することでがん細胞の死滅を誘導できるわけです。当然のことながらこの場合のゴールは抗がん剤の開発になります。まとめると、生体の反応を分子レベルで理解することで重要な分子間相互作用箇所が明らかとなります。その特定箇所を標的とした新規の化合物の開発を行えば、新規の抑制法の開発に繋がり、応用次第では産業化の糸口を作りだすことに貢献できるということです。このような過程を辿ることにより、私は新規薬剤の創製や新規生体反応制御因子の創製を目指します。
7. 独り言 上手くありませんがPSPでたまに将棋で遊んでいます。レベルをだんだんと上げていくと、負けが込んできます。負ける度に思うのですが、もう少し先を読めることができれば勝てるのではないかということです。研究においても、目先のことしか考えていなくて、後になってはっと気がつくことがしばしばです。生物を研究対象にしていると、まだまだ明らかになっていなことは多く、全く先が読めない状況にあることを痛感しています。 転写反応をメインに行っていた2000年頃までは、「遺伝子発現の活性化=転写の活性化」だと信じていました。信じていたというところが重要です。そうです、単純にそう思っていたことに他なりません。もう少し付け加えるならば、遺伝子発現の活性化制御研究はmRNA量が増える反応の解析のみで解決できると単純に考えていました。しかし、2000年以降に着手したゼブラフィッシュの初期胚研究で気がつきました。すなわち、mRNA供給なしにタンパク質量が増加する遺伝子発現制御を知る(認識する)こととなったわけです。ここでいい訳ですが、転写研究真っ盛りという時代が私にそう信じ込ませたのかもしれません。視野が狭かった、考えが浅かった、いずれにしても翻訳反応の活性化に目が向くようになってきました。 遺伝子発現を解析する際に、数多くの論文ではRT-PCRでmRNA量を間接的に測定して遺伝子発現レベルとしています。しかし、母性mRNAによる制御を鑑みるとこれは正しくないと言えます。すなわち、見掛け上mRNA量の変化はなくても質的な変化、すなわちpoly(A)鎖伸長の変化で翻訳の活性化が誘導され、結果的にタンパク質量の増加がもたらされるという現象を考慮に入れることが重要と思うからです。 そう考えると、最近、私はマイクロRNA(miRNA)による遺伝子発現制御メカニズムに興味が湧いてきました。miRNAはmRNAの3'UTRに作用して、poly(A)鎖の短小化(deadenylation)を促進し、結果的に翻訳反応を抑制します。また、この抑制はmRNAの分解促進を伴わないことが明らかとなってきました。すなわち、mRNAの減少なしに遺伝子発現レベルは低下するわけです。今後は、miRNAによる発現制御メカニズムも視野に入れて研究を進めていきたいと考えています。 生命に関わる諸反応は化学反応です。よって、必ず分子が関与し、化学の法則が適応されます。私は学会や班会議で何度か「あなたの解析している分子は1つの細胞に何個存在しているのですか」と質問したことがあります。残念ながらこの質問にその場で的確に答えられた演者はほとんど記憶にありません。なぜ、生命系の研究者は分子数をあまり考慮しないのでしょうか?私にとっては不思議です。持論(と言うより、恩師の半田教授の影響も大)ですが、分子数で反応を考えるべきだと思っています。アボガドロ数を6.02X10の23乗と覚えていれば、細胞あたりの分子数は簡単に計算できます。 「柳の木の下に二匹目のドジョウはいない」と言われます。要するに、同じことをやっても、そうそう簡単には成功しないことだと私は解釈しています。 私の実験では先に説明しました通り転写制御研究で低分子化合物DRBを分子プローブとして使い、試験管内転写実験において新規転写伸長因子の同定・精製および生化学的・分子生物学的解析を行いました。その結果、動物細胞における転写伸長段階の一時停止メカニズムを明らかにすることができました。そして、今後は同じ手法を翻訳制御研究に持ち込み、すなわち低分子化合物で試験管内翻訳反応を阻害することによって新規制御因子や最終的には新規のメカニズムを明らかにできるのではないかと考えています。まさに、柳の下の二匹目のドジョウを狙っているわけです。
8. 余談 −影響を受けた著書 ・7つの習慣(スティーブン・R・コヴィー著、キングベアー出版) この著書から学んだことはここで書ききれないほどたくさんあるのですが、強いて抽出すると、相互依存、すなわちお互いに良いところを出し合うことで仕事を推進する有益性、第二の領域、すなわち健康維持や体力増進・友好関係の樹立・新しい分野の勉強・集中力を増すための環境作りなどが重要である点、さらには委譲 (delegation)によって 自分以外の人に仕事を任せる事で自分が苦手(いやでやりたくない)な仕事やストレスが溜まる仕事を他人との協力で克服することの必要性です。何度も読み返せる素晴らしい本です。 ・さあ才能に目覚めよう(マーカス・バッキンガム&ドナルド・O・クリフトン著、日本経済新聞出版社) それぞれの得意分野で勝負することを目的として書かれたこの本は、自分の得意分野や長所を見出す助けとなります。これによって自分の能力を意識し、さらに仲間との関係の確立でよりよい仕事環境が整うのではないでしょうか? この著書も、何度読んでも学ぶことばかりです。 ・減らす技術 The Power of LESS(レオ・バボータ著、ディスカヴァー・トゥエンティワン) この著書からは、時間の使い方を学びました。渋滞に巻き込まれないようにするためには、多くの人と異なるタイムテーブルで行動するメリット、朝早く起きることだけで生活が一変する事実など、無駄な時間をなくす必要性を痛感しました。是非、読んでみてください。 Kさんは100mを11秒台で走れます。Hさんは400mを40秒台で走ることができます。両人とも、きわめて立派な記録で鍛えていない人には到底出来ることではありません。私は、残念ながらそのような高速で走ることはできません。例え練習したとしても無理です。一方、両人はマラソンクラスの40kmとなると、これをある程度の速度で走ることができないのです。正確に言うと、走ろうとは思わないのです。ここで私を仮にTさんと呼ぶことにします。Tさんは100mや400mを速く走れるようになりたいのですが、先ほども書きましたように練習しても一向に速く走れません。しかし、40kmだったら最終的には時速10km/hで走ることができます。 注:*東京工業大学統合研究院教授半田宏先生の研究室での研究成果 **現在は第一三共株式会社 開発第一部第1G 所属、 ***現在は東京工業大学大学院生命理工学研究科准教授。
|